Auditory cortex: physiology (Norsk)
De anatomiske og funksjonelle egenskapene til den menneskelige auditive cortex er svært komplekse, og mange spørsmål gjenstår fortsatt om integreringen av auditiv informasjon på dette nivået.
Historie
De første studiene som forbinder strukturen og funksjonen til hjernebarken i den temporale lappen med auditiv persepsjon og talen ble utført av Paul Broca (1824-1880) og Carl Wernicke (1848-1904). Beskrivelser av Brocas afasi (taleforstyrrelse forårsaket av en lesjon i Brodmans områder 44 og 45 – også kjent som Brocas område) og Wernickes afasi (en forstyrrelse i taleoppfatning forårsaket av skade på Brodmans område 22) gjorde det mulig å bestemme plasseringen av hørsel og talebehandling i hjernebarken.
Funksjonen til den auditive cortexen
Klassisk har to hovedfunksjonelle regioner blitt beskrevet i auditiv cortex:
- Primær auditiv cortex (AI), sammensatt av nevroner som er involvert i dekoding av cochleotopisk og tonotopisk romlig representasjon av en stimulus.
- Sekundær auditiv cortex (AII), som ikke «t har tydelig tonotopisk organisering, men har en viktig rolle i lydlokalisering og analyse av komplekse lyder: spesielt for spesifikke dyrevokaliseringer og menneskespråk. Den har også en rolle i hørselsminnet.
- Belteområdet, rundt AI og AII, som hjelper til med å integrere hørsel med andre sensoriske systemer.
Funksjonen til den primære auditive cortex
I AI er nevroner selektive for bestemte frekvenser og er ordnet i isofrekvensbånd som er tonotopisk organisert. Den nøyaktige romlige fordelingen av isofrekvensbåndene er relatert til organiseringen av hørselsreseptorene. Aktiviteten deres avhenger av stimulansens egenskaper: frekvens, intensitet og plassering av lydkilden i rommet. Funksjonelt er denne regionen sterkt påvirket av motivets våkne tilstand. En rekke meget spesifikke nevroner i AI er også involvert i analysen av komplekse lyder.
Nye teknikker for å studere hjernebarken (funksjonell magnetisk resonansavbildning: fMRI; positronemisjonstomografi: PET; og magnetoencefalografi: MEG ) antyder at frekvensfordelingen sett hos dyr (med tradisjonelle eksperimentelle metoder) ikke samsvarer nøyaktig med den som er sett hos mennesker, selv om de alle har isofrekvensbånd, sett med MEG nedenfor. fMRI hos mennesker antyder at lave frekvenser er kodet i de overfladiske posterolaterale områdene i sylvissprøyten, mens høye frekvenser ligger i de dypere og anteromediale områdene. Det er imidlertid viktig å merke seg at det eksisterer en viss variasjon mellom individer.
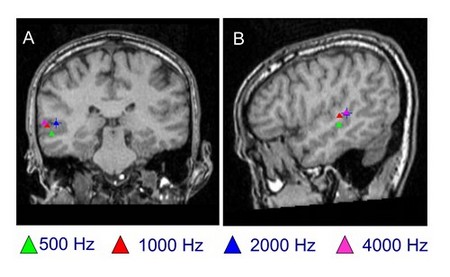 |
Magnetenoencefalografi (MEG): lokalisering av rene toner i et normalt hørselsperspektiv Lokalisering av rene toner (500 Hz, 1000 Hz, 2000 Hz og 4000 Hz) i frontal (A) og laterale (B) plan. Image P. Gil-Loyzaga, Center MEG de l «Université Complutense (Madrid). |
Temporal integrasjon av auditive stimuli
Når de er våkne, er mennesker, i likhet med andre dyr, i stand til å oppfatte de små tidsmessige variasjonene av komplekse lyder. variasjoner er avgjørende for forståelsen av menneskelig tale. En rekke studier som undersøker AI har identifisert at i våken primater, to forskjellige populasjoner av henholdsvis synkrone og asynkrone nevroner koder for sekvens ial stimuli annerledes.
- Synkrone nevroner analyserer langsomme tidsendringer. De reagerer nøyaktig på lavfrekvent stimulering (A1), men klarer ikke å opprettholde aktiviteten hvis antall stimuli øker. De raske hastighetsendringene oppfattes av disse nevronene som en kontinuerlig tone. De er involvert i både frekvens- og intensitetsanalyse.
- Asynkrone nevroner analyserer raske tidsendringer (av mange stimuli). De kan bestemme variasjoner i kort varighet og kan skille nøyaktig en stimulans fra den neste.
Den funksjonelle inndelingen av den auditive cortex gjør det mulig å avkode tidsvariasjoner av en stimulus ekstremt nøyaktig sammenlignet med andre sentre av hørselsveien. Det gjør det mulig å få mer informasjon om komplekse lyder, samt plasseringen av en lydkilde og dens bevegelse.
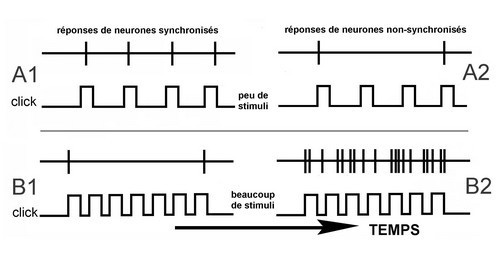
Figur: Tilbakemelding des neurones synchronises – > Respons i synkrone celler
Reponses des neurones non-synchronises – > Respons i asynkrone celler
Peu de stimuli – > få stimuli
Beaucoup de stimuli – > mange stimuli
Temps – > Time
Synchronous and asynchronous neurons
- Synchronous neurons alltid svare på hver stimulus (klikk) når stimulustogene har intervaller større enn 20 ms (A1). Etter hvert som intertreningsintervallet synker (dvs. repetisjonshastigheten blir raskere), begynner disse nevronene å desychronisere skytefrekvensen. Når interstimulusintervallet faller under 10 ms (B1), skyter disse nevronene bare i begynnelsen og slutten av stimulusen (henholdsvis debut og offsetrespons).
- Asynkrone nevroner reagerer ikke synkront på stimuli ( A2 og B2), men deres aktivitet øker gradvis til en veldig høy utslippshastighet (B2).
Spektral integrasjon av auditiv stimuli
Dyresang og menneskespråk varierer sterkt mellom individer. Frivillige og ufrivillige variasjoner eksisterer også innenfor samme emne. Selv om oppfatningen av hørselsmeldinger krever analyse av frekvensene som utgjør en kompleks lyd, er spektralanalyse enda viktigere.
Hvis lydspekteret inneholder hele lydbølgeprofilen til en kompleks lyd (lydkonvolutten) opprettholdes, kan god hørsel og fonemforståelse oppstå, selv når visse spesifikke frekvenser fjernes.
Ikke-invasiv MEG-avbildning kan brukes til å nøyaktig bestemme med utmerket romlig presisjon, plasseringen av fremkalt aktivitet som oppstår i noen få millisekunder. MEG er en passende teknikk for å studere kompleks auditiv funksjon, for eksempel tale, samt potensielle funksjonelle effekter av kortikal skade.
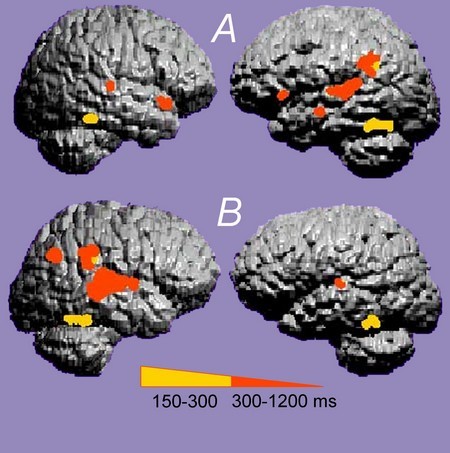 |
MEG i et normalt emne (A) og et dyslektisk emne (B) Hos personer med normal hørsel (A), spesifikk språklig kortikal aktivering forekommer hovedsakelig i den venstre auditive cortex. Hos dyslektiske forsøkspersoner (B) er aktivering mer fremtredende i høyre cortex og er mer diffus. Image P. Gil-Loyzaga, Center MEG de l «Université Complutense (Madrid) |