Fronteiras na microbiologia celular e de infecções
Introdução
Staphylococcus aureus é um importante patógeno humano isolado de pacientes hospitalizados em todo o mundo, que causa infecções hospitalares e adquiridas na comunidade ( Lowy, 1998). Esse patógeno é o agente etiológico de diversas infecções sistêmicas, afetando pele e tecidos moles, bem como sistemas musculoesquelético e circulatório (Lowy, 1998; Changchien et al., 2016). Foi relatado que S. aureus pode sobreviver em macrófagos derivados de monócitos humanos (Kubica et al., 2008). A virulência de S. aureus está intimamente associada a uma variedade de enzimas secretadas e toxinas produzidas pela bactéria (Otto, 2014). Hemolisina, leucocidina (Panton-Valentine leucocidina, PVL) e toxina-1 da síndrome do choque tóxico (TSST-1), facilitando o dano à membrana dos glóbulos vermelhos, prejudicando a função fagocítica dos leucócitos e induzindo a síndrome do choque tóxico, respectivamente, são essenciais para o processos patogênicos de S. aureus (Löffler et al., 2010; Vandenesch et al., 2012; Berube e Bubeck Wardenburg, 2013; Andrey et al., 2015; Al Laham et al., 2015). Estudos recentes demonstraram que a hemolisina também participa da formação do biofilme de S. aureus (den Reijer et al., 2016). O aumento da prevalência de cepas de S. aureus multirresistentes em ambientes hospitalares e comunitários aumenta ainda mais os perigos de S. aureus e representa um sério desafio para a terapia clínica (Voss e Doebbeling, 1995; Evangelista Sde e de Oliveira, 2015).
Recentemente, várias cepas pertencentes a uma classe de S. aureus com um fenótipo hemolítico incompleto (SIHP) foram encontradas em nosso hospital. A hemólise causada por essas cepas de SIHP é significativamente diferente do anel hemolítico completo (fenótipo β-hemolítico) produzido em outras cepas de S. aureus. No entanto, essas cepas de SIHP ainda não foram identificadas e caracterizadas de forma abrangente. Para explorar as características microbiológicas dessas cepas de SIHP, coletamos 60 cepas de SIHP e as estudamos usando vários critérios, incluindo fenótipo hemolítico, expressão do gene da hemolisina, características de resistência a drogas e virulência. Este estudo demonstra que SIHPs são cepas resistentes à meticilina que expressam altamente β-hemolisina e possuem um alto potencial de virulência.
Materiais e métodos
Cepas bacterianas
Sessenta SIHP as cepas foram isoladas de pacientes internados no segundo hospital afiliado da Soochow University entre 2013 e 2015. Amostras duplicadas de cada paciente foram coletadas para análise. Esses isolados foram então cultivados em placas de ágar sangue de carneiro Columbia (CHROMagar Company, Shanghai, China) a 35 ° C em uma atmosfera contendo 5% de CO2 (v / v). A identidade das cepas como S. aureus foi confirmada usando o sistema de microbiologia automatizado Phoenix-100 (Becton, Dickinson and Company, EUA). As cepas de controle de S. aureus, com fenótipo hemolítico completo, foram isoladas de pacientes em nosso hospital durante o mesmo período. Aqui, S. aureus com o fenótipo hemolítico completo foi chamado de cepas de SCHP. A cepa de referência S. aureus ATCC25923 (Shanghai Center for Clinical Laboratory, China) tem o fenótipo hemolítico completo e também atuou como cepa de controle. O Comitê de Ética Médica do Segundo Hospital Afiliado da Soochow University aprovou este estudo e todos os isolados foram coletados com o consentimento do paciente neste estudo.
Análises comparativas do fenótipo hemolítico incompleto
O S. aureus as cepas foram cultivadas em placas comerciais de ágar sangue de diferentes empresas (CHROMagar, Autobio Diagnostics Co., Ltd., China e BioMérieux, China), bem como placas de ágar sangue de ovelha auto-preparadas usando ágar sangue Columbia em pó (OXIDE, Reino Unido). As bactérias foram cultivadas a 35 ° C em atmosfera contendo 5% de CO2 (v / v) por 24 he, em seguida, passaram por passagens seriadas. O fenômeno hemolítico foi então observado. As cepas clinicamente isoladas com fenótipo hemolítico completo e a cepa de referência ATCC25923 também foram observadas como controle para análises comparativas.
Determinação dos níveis de mRNA de quatro genes de hemolisina de S. aureus usando transcrição reversa quantitativa em tempo real PCR (qPCR)
A extração de RNA total do SIHP e da cepa de referência ATCC25923 foi realizada conforme descrito anteriormente (Qin et al., 2014). Inicialmente, as células foram lisadas usando lisostafina, lisozima e proteinase K. O RNA foi então extraído e purificado usando RNeasy Mini Kit (Qiagen) de acordo com o protocolo recomendado pelo fabricante. A qualidade e a concentração do RNA foram avaliadas usando o NanoDrop1000. O RNA foi então reverso -transcrito para cDNA usando um kit de transcrição reversa (Thermo Fisher Scientific Inc., EUA) de acordo com as instruções do fabricante.Resumidamente qPCR foi conduzido da seguinte forma: pré-desnaturação a 94 ° C por 3 min, desnaturação a 94 ° C por 30 s, recozimento a 52 ° C por 30 s, alongamento a 72 ° C por 40 s, um total de 45 ciclos . Cada amostra teve três repetições técnicas e três biológicas. O perfil transcricional de cada um dos quatro genes da hemolisina e do gene 16s rRNA foi determinado pelo método 2 − ΔΔCt. O nível de transcrição foi determinado em relação à expressão do gene 16s rRNA. As sequências de quatro genes de hemolisina (hla, hlb, hlgC e hld) foram recuperadas do banco de dados GenBank. O software Primer 5.0 foi usado para projetar os primers (Tabela 1) e esses primers foram sintetizados por Sangon Biotech Co., Ltd. (Shanghai, China).
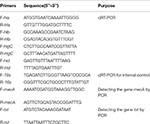
Tabela 1. Primers usados neste estudo.
SPSS 17.0 foi usado para análise de dados. Os dados de medição são apresentados como x̄ ± s. O teste T para duas amostras independentes foi usado para comparar a expressão relativa de cada um dos quatro genes da hemolisina. A significância estatística foi definida como p < 0,05.
Detecção da expressão de α-hemolisina (Hla) por Western Blot
A concentração de O SIHP e a cepa de referência S. aureus ATCC25923 foram ajustados para 5,0 McFarland usando um medidor de turbidez. A proteína total foi então extraída de cada amostra, ajustada para a mesma concentração e submetida a eletroforese em gel SDS-PAGE. As proteínas foram então transferidas para uma membrana de nitrocelulose e a membrana foi bloqueada com leite desnatado a 5% por 1 h. Anticorpos policlonais de α-hemolisina estafilocócicos de cabra (Abcam) foram adicionados em uma concentração final de 2 μg / mL. A mistura de reação foi então incubada a 4 ° C durante a noite. Após a membrana ser lavada, anticorpos IgG anti-cabra de coelho marcados com HRP diluídos 1: 1000 foram adicionados e incubados a 37 ° C durante 2 h. Finalmente, substratos quimioluminescentes foram adicionados para o desenvolvimento da cor. A expressão de α-hemolisina foi finalmente observada sob o imageador.
Teste de sensibilidade antimicrobiana de SIHP
O método de diluição em caldo de microtitulação foi usado para realizar a triagem de susceptibilidade antimicrobiana. O procedimento foi realizado de acordo com o manual operacional do sistema automatizado de microbiologia Phoenix-100. Os resultados desta triagem foram interpretados usando os critérios M100-S24 introduzidos pelo Clinical and Laboratory Standards Institute (CLSI; Clinical and Laboratory Standards Institute, 2014). O status de S. aureus resistente à meticilina (MRSA) foi determinado usando a MIC (concentração inibitória mínima) de dois antibióticos. Para as cepas SIHP ≥4 μg / mL foi o MIC para oxallicina, e ≥8 μg / mL foi o MIC para cefoxitina.
Detecção dos genes mecA e tst
Todos os 60 SIHP as cepas foram testadas para o gene mecA de resistência a drogas e o gene de virulência tst. A extração por fervura do DNA genômico foi conduzida após a lise celular usando lisostafina. Os genes mecA e tst foram então amplificados a partir do DNA genômico usando o kit DreamGreen Taq. As condições de PCR foram as seguintes: pré-desnaturação a 94 ° C durante 3 min, desnaturação a 94 ° C durante 30 s, hibridação a 50 ° C durante 30 s, extensão a 72 ° C durante 1 min. Os produtos de PCR foram sequenciados e analisados pelo BLAST para validar como sendo os produtos esperados. As sequências do gene mecA de resistência a drogas e do gene de virulência tst de S. aureus foram recuperadas do banco de dados do GenBank. O software Primer 5.0 foi usado para projetar os primers (Tabela 1) e esses primers foram sintetizados por Sangon Biotech Co., Ltd. (Shanghai, China).
Detecção da sobrevivência de cepas SIHP de S. aureus em Macrófagos
Para comparar a sobrevivência de cepas de S. aureus SIHP em macrófagos com a de cepas de S. aureus SCHP tst negativas, a linha de células de monócitos humanos THP-1 foi mantida em RPMI-1640 contendo 10% (v / v) FBS a 37 ° C em uma atmosfera contendo 5% (v / v) de CO2. Para a infecção de macrófagos, células THP-1 foram semeadas a 5 × 105 células por poço em placas de cultura de tecidos de 24 poços e induzidas a se diferenciar por 10−7 M de forbol 12-miristato 13-acetato (PMA) por 48 h. Aproximadamente 3 × 108 unidades formadoras de colônia (CFU) de bactérias de fase logarítmica (OD600 0,5–0,6) foram sedimentadas por centrifugação, lavadas duas vezes com PBS e ressuspensas em 1 ml de RPMI-1640. Em seguida, as bactérias foram adicionadas à monocamada de células a um MOI de 20: 1 e centrifugadas por 5 min a 1000 rpm. As células infectadas foram incubadas por 20 min a 37 ° C, então foram lavadas três vezes com PBS pré-aquecido (pH 7,4) e incubadas por mais 1 h em meio contendo 100μg / ml de gentamicina para matar bactérias extracelulares. As células foram lavadas e lisadas para quantificar a bactéria intracelular como a amostra de tempo zero (T0). Células adicionais foram coletadas após 12 ou 24 horas de incubação na presença de meio de cultura de tecido suplementado fresco contendo 12 μg / ml de gentamicina.O crescimento de bactérias em células THP-1 foi determinado dividindo o número de bactérias intracelulares em 12 h ou 24 h pelo número no tempo 0 (T12 / T0 ou T24 / T0). O experimento foi repetido independentemente três vezes.
Detecção de secreção de citocinas de macrófagos infectados por cepas de S. aureus SIHP
Os sobrenadantes de cultura de macrófagos THP-1 infectados por 12 h por bactérias foram coletados. Os níveis de citocinas foram medidos por citometria de fluxo FACSCalibur (BD) usando um CBA Human Thl / Th2 Cytokine Kit II (BD) de acordo com as instruções do fabricante.
Multilocus Sequence Typing and Analysis
Resultados
Fenótipo hemolítico de cepas SIHP em placas de sangue de ágar de ovelha
Como mostrado nas Figuras 1A, B, o anel hemolítico completo (fenótipo β-hemolítico) foi observado no controle S . cepas de aureus após cultura de cepas em placas de ágar sangue por 24 h. No entanto, diferentes fenótipos hemolíticos (chamados de fenótipo hemolítico incompleto aqui) foram exibidos em cepas de SIHP. Além disso, o fenótipo de hemolítico incompleto pode ser observado nas cepas de SIHP que cresceram em diferentes ambientes condições incluindo condições microaerofílicas, aeróbicas e anaeróbicas, respectivamente (Figuras 1C-E). Além disso, após 10 passagens em série, este fenótipo hemolítico incompleto ainda era mantido (Figura 1F).
Relativa Níveis de expressão de mRNA de quatro genes de hemolisina
Conforme mostrado na Figura 2, os níveis de expressão dos quatro genes de hemolisina hla, hlb, hlgC e hld em cepas de SCHP não exibiram diferenças estatisticamente significativas (p > 0,05) em comparação com as cepas de controle ATCC25923. No entanto, os níveis de expressão de hla, hlgC e hld em cepas de SIHP foram significativamente suprimidos por 50, 16,7 e 8,3 vezes respectivamente (p < 0,05) em comparação com o de cepas de controle ATCC25923 e cepas de SCHP, enquanto a expressão de hlb em cepas de SIHP foi significativamente aumentada 7,7 vezes em comparação com os controles (p < 0,05).
Detecção de α-hemolisina por Western Blot
A α-hemolisina codificada por o gene hla é uma toxina formadora de poros secretada por S. aureus e seu peso molecular é 33 kDa (Andrey et al., 2015; Al Laham et al., 2015). A análise de Western blot mostrou que a expressão de α-hemolisina em cepas de SIHP foi 40 vezes menor do que a de SCHP e cepas de controle, o que é consistente com os dados de qRT-PCR (Figura 3).

Figura 3. Western blot de α-hemolisina em S. aureus. ATCC29213: controle de S. aureus com o fenótipo hemolítico completo; SCHP: S. aureus com o fenótipo hemolítico completo isolado de amostras clínicas no mesmo período; SIHP: S. aureus com um fenótipo hemolítico incompleto.
Resistência a medicamentos de SIHP
Testada a suscetibilidade aos medicamentos usando diluições de caldo de microtitulação demonstrou que os valores de MIC para oxacilina e cefoxitina foram todos > 2 e > 8 μg / mL, respectivamente, em 60 manchas de SIHP . De acordo com as diretrizes do CLSI M100-S24, todas as 60 cepas de SIHP são classificadas como cepas de MRSA. A PCR também detectou o gene mecA em todas as 60 cepas.
Teste para o gene tst
O gene tst é um importante fator de virulência em S. aureus. A PCR mostrou que todas as 50 cepas SIHP carregavam genes tst.
A capacidade de sobrevivência de cepas SIHP de S. aureus em macrófagos é maior do que a de S. aureus tst negativo
Nós comparamos as habilidades de sobrevivência intracelular de cepas de SIHP e cepa de SCHP tst negativo em macrófagos. Ao contar o número de bactérias recuperadas das placas, descobrimos que a capacidade de sobrevivência intracelular da cepa SIHP era muito maior do que a da cepa de S. aureus tst negativa após infectados macrófagos derivados de THP-1 12 ou 24 h (Figura 4). Preliminarmente, especulamos que as cepas de SIHP possuem um potencial de alta virulência, e foi sugerido que o SIHP deveria receber mais atenção no hospital.
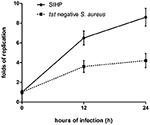
Figura 4. Sobrevivência de cepas SIHP e S. aureus tst negativo em macrófagos derivados de THP-1. Após os macrófagos serem infectados por bactérias por 12 e 24 h, os macrófagos foram lisados e as bactérias sobreviventes intracelulares foram semeadas em ágar LB. As dobras de replicação de bactérias sobreviventes intracelulares foram calculadas de acordo com o número de colônias na placa de ágar LB acima.
As diferenças de secreção de citocinas e quimiocinas de macrófagos infectados por cepas SIHP de S. aureus e S. aureus tst negativo
Para comparar as diferenças de secreção de citocinas e quimiocinas em macrófagos derivados de THP-1 infectados pela cepa SIHP e S. aureus tst negativo.cepa aureus, detectamos citocinas pró-inflamatórias por citometria de fluxo no sobrenadante da cultura de macrófagos infectados por cepas SIHP e cepa tst negativa de S. aureus. Os resultados mostraram indução significativamente maior de IL-2, IL-6 e IL-17A em cepas SIHP, mas não por S. aureus tst negativo, sugerindo que cepas SIHP induzem respostas de citocina / quimiocina nos macrófagos (Figura 5). As outras citocinas IL-4, IL-10, INF-γ e TNF secretadas por macrófagos não apresentaram diferenças óbvias entre as cepas SIHP e os grupos afetados de S. aureus tst negativo.
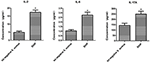
Figura 5. Detecção de citocinas e quimiocinas secretadas por macrófagos infectados com S. aureus. * P < 0,05.
Tipagem de sequência multifoco
Determinamos o genótipo multilocus de 60 isolados de SIHP coletados de pacientes internados em nosso hospital entre 2013 e 2014. O esquema de sete locus recomendado no banco de dados MLST de S. aureus foi aplicado. A análise de MLST mostrou que todas as cepas de SIHP foram classificadas no clone ST5.
Discussão
A hemolisina é um dos fatores de virulência mais importantes para S. aureus (Wiseman, 1975). Os efeitos combinados de cada um dos quatro tipos de hemolisina resultam na destruição da membrana dos glóbulos vermelhos, o que leva à formação do anel hemolítico transparente completo nas placas de ágar sangue. Nos últimos anos, o SIHP foi encontrado nas amostras clínicas do segundo hospital afiliado da Soochow University. Neste estudo, buscou-se identificar essas cepas e estudar suas características em microbiologia. Inicialmente, usamos diferentes placas de ágar sangue de carneiro comerciais ou auto-preparadas para isolar, incubar e passar em série as cepas. Mostramos que existem algumas cepas que exibem e mantêm um fenótipo hemolítico incompleto mesmo após subcultura prolongada. Assim, a possibilidade de que esse fenótipo seja induzido por fatores não bacterianos pode ser excluída. Nossos estudos sugerem que o SIHP pode ser um subconjunto de cepas únicas de S. aureus.
Estudos anteriores sugeriram que os glóbulos vermelhos de coelho são altamente sensíveis à α-hemolisina (Hildebrand et al., 1991). Berube et al. relataram que o efeito da α-hemolisina na lise dos glóbulos vermelhos é dependente da concentração (Berube e Bubeck Wardenburg, 2013). A γ-hemolisina pode danificar as células vermelhas do sangue de humanos e animais (Kaneko e Kamio, 2004). Enquanto a δ-hemolisina danifica os glóbulos vermelhos, apenas em altas concentrações, ela forma um poro transmembranar que causa a lise da membrana celular (Verdon et al., 2009). Neste estudo, mostramos que a expressão de quatro hemolisinas de cepas de SIHP e cepas de SCHP tem perfis de expressão transcricional significativamente diferentes. A expressão de α-, γ-, δ-hemolisina nas cepas de SIHP, que poderiam danificar as células vermelhas diretamente, é muito menor do que as cepas de SCHP, enquanto a β-hemolisina tem níveis de expressão muito mais elevados em comparação com as cepas de SCHP. Além disso, o nível de expressão da proteína de α-hemolisina foi posteriormente validado por Western blot. Foi relatado que a β-hemolisina aumenta predominantemente a sensibilidade dos glóbulos vermelhos a outras toxinas em vez de lisar as células diretamente, a menos que as células sejam cultivadas em temperaturas mais baixas (Vandenesch et al., 2012). Neste estudo, a alta expressão de β-hemolisina foi detectada nas cepas de SIHP, mas o anel hemolítico completo nunca foi observado nas cepas de SIHP, mesmo quando incubadas a 4 ° C. Estes resultados sugerem que o fenótipo incompleto observado de cepas de SIHP é mais provavelmente os efeitos de todas as quatro hemolisinas que são expressas de uma maneira diferente em comparação com as cepas de SCHP. Estudos anteriores demonstraram que durante a patogênese crônica, o efeito combinado de pressões seletivas resultantes de antibióticos, incluindo ciprofloxacina e trimetoprim, bem como a resposta imune do hospedeiro, pode levar ao aumento da expressão de α-hemolisina, que é muito importante para a colonização de S .aureus na mucosa, especialmente em infecções do trato respiratório (Goerke et al., 2006; Huseby et al., 2010). Neste estudo, todos os pacientes infectados com SIHP receberam tratamento de longo prazo com antibióticos de amplo espectro, se esta pressão seletiva induziu a superexpressão de genes resistentes a medicamentos e interferiu na expressão dos genes da hemolisina permanece uma possibilidade que precisa ser mais detalhada exame.
Para explorar as características microbiológicas do SIHP, conduzimos testes de suscetibilidade aos antimicrobianos e escolhemos o gene de virulência tst que está associado à patogenicidade do S. aureus. Descobrimos que todas as 60 cepas de SIHP podem ser classificadas como cepas de MRSA e carregam o gene mecA. Os resultados dos testes de sensibilidade aos antimicrobianos mostraram que mesmo após 10 passagens, dessas 60 cepas de SIHP, os valores de MIC de linezolida e teicoplanina de apenas três cepas de SIHP mudaram pouco.No entanto, essas mudanças nos valores de MIC não influenciaram a avaliação dos resultados de suscetibilidade aos antimicrobianos. O gene de virulência tst codifica a toxina da síndrome do choque tóxico (TSST-1) (Kreiswirth, 1989). Como um superantígeno, o TSST-1 aumenta as respostas de choque e imunossupressão induzidas por endotoxinas (Kulhankova et al., 2014). O teste de PCR descobriu que todas as 60 cepas de SIHP possuíam o gene tst, sugerindo aumento da virulência. Recentemente, muitos relatórios mostraram que S. aureus poderia sobreviver em macrófagos derivados de monócitos humanos e essas cepas de S. aureus são mais virulentas (Kubica et al., 2008; Tranchemontagne et al., 2015; Münzenmayer et al., 2016; Nandi e Bishayi, 2016). Portanto, comparamos as habilidades de sobrevivência intracelular de cepas de SIHP e cepas de SCHP que é tst negativo de S. aureus em macrófagos, e descobrimos que a capacidade de sobrevivência intracelular de cepa de SIHP foi muito maior do que a cepa de S. aureus tst negativo. Além disso, a análise dos dados do paciente mostrou que o efeito da terapia anti-infecciosa na maioria dos pacientes infectados com cepas de SIHP foi pior do que em pacientes semelhantes infectados com cepas de SCHP (dados não mostrados). Nosso estudo fornece evidências experimentais preliminares para a especulação de que as cepas de SIHP possuem um potencial de alta virulência.
Durante o processo de infecção de S. aureus invasivo, as moléculas associadas ao patógeno iniciam o sistema imunológico inato levando à ativação e recrutamento de neutrófilos e macrófagos e a produção de citocinas pró-inflamatórias, mais notavelmente TNF-α, IL-1β e IL-6 (Bekeredjian-Ding et al., 2015; Giai et al., 2016; Zhao et al., 2016 ) Foi relatado que a secreção de citocinas inflamatórias IL-6 aumentou com o aumento das concentrações de S. aureus (Chen et al., 2016). Além disso, os S. aureus superantigênicos são particularmente eficientes na estimulação da produção de IL-17 (Islander et al., 2010). Neste estudo, as cepas de SIHP mostraram indução significativamente maior de IL-2, IL-6 e IL-17A de macrófagos em comparação com cepas de SCHP tst negativas, o que sugere que as cepas de SIHP poderiam induzir mais secreção de citocinas contra o efeito de matar de macrófagos. Além disso, a análise de MLST mostrou que todas as cepas de SIHP foram classificadas no clone ST5. Além disso, essas cepas de SIHP foram agrupadas no tipo spa de t2460 (dados não mostrados).
Em conjunto, o SIHP pode ser um novo subconjunto de MRSA com potencial alta virulência, e mais atenção deve ser dada em controle e tratamento dessas cepas em hospitais.
Contribuições dos autores
Declaração de conflito de interesses
Os autores declaram que a pesquisa foi conduzida na ausência de qualquer comercial ou relações financeiras que podem ser interpretadas como um potencial conflito de interesses.
Agradecimentos
Este estudo foi financiado pela Fundação Nacional de Ciências Naturais da China (81572032, 81401636), a Ciência Natural Fundação para Faculdades e Universidades na província de Jiangsu (16KJB320006) e o Programa de Ciência e Tecnologia de Suzhou (SYSD2014094, SYS201551, SS201638).
Clínica e Laboratório Standards Institute (2014). Padrões de desempenho M100-S24 para testes de susceptibilidade antimicrobiana; Vigésimo quarto suplemento informativo. Wayne, PA: CLSI.
Nandi, A., e Bishayi, B. (2016). Os Staphylococcus aureus sobrevividos intracelularmente após a fagocitose são mais virulentos na indução de citotoxicidade em macrófagos peritoneais murinos frescos utilizando TLR-2 como um possível alvo. Microb. Pathog. 97, 131–147. doi: 10.1016 / j.micpath.2016.06.007
Resumo PubMed | CrossRef Full Text | Google Scholar
Wiseman, G. M. (1975). As hemolisinas de Staphylococcus aureus. Bacteriol. Rev. 39, 317–344.
PubMed Resumo | Google Scholar